 Return
to Genesis of Eden?
Return
to Genesis of Eden? Fractal Neurodyamics and
Quantum Chaos :
Fractal Neurodyamics and
Quantum Chaos :
Resolving the Mind-Brain Paradox Through Novel Biophysics
Chris King Mathematics and Statistics Department University of Auckland
in Mac Cormac Earl, Stamenov Maxin (eds)
Fractals Of Brain, Fractals of Mind , Advances in Consciousness Research
7, John Betjamins 179-232.
ISBN 90-272-5127-4 (Eur) ISBN 1-55619-187-1 (US)
Abstract: A model of the mind-brain relationship is developed in which novel biophysical principles in brain function generate a dynamic possessing attributes consistent with consciousness and free-will. The model invokes a fractal link between neurodynamical chaos and quantum uncertainty. Transactional wave collapse allows this link to be utilized predictively by the excitable cell, in a way which bypasses and complements formal computation. The formal unpredictability of the model allows mind to interact upon the brain, the predictivity of consciousness in survival strategies being selected as a trait by organismic evolution.
1 Mind and Brain, Chaos and Quantum Mechanics.
1.1 Paradigms in Scientific Discovery and The Enigma of Consciousness The twentieth century has seen the unification of the microscopic and cosmic realms of physics in theories such as inflation, in which symmetry-breaking of the fundamental forces is linked to cosmic expansion. Molecular biology has had equally epoch-making successes unravelling the intricate molecular mechanisms underlying living systems, from the genetic code through to developmental structures such as homeotic genes. Despite these conceptual advances, the principles by which the brain generates mind remain mysterious. The intractability of this central unresolved problem in science suggests its principles run deeper than the conventional biochemical description, requiring novel biophysical principles. This paper develops such a model based on linkage between the fractal aspect of chaotic neurodynamics and quantum non-locality, giving brain science a cosmological status at the foundations of physical description.
1.2 Chaos and Quantum Mechanics : Unpredictability as a Basis for Mind Although mechanists tolerate mind as a passive shadow representation of physical causality in such theories as epiphenomenalism and brain-mind identity, free-will conjures up the spectre of active causal interference. To quote Sir John Eccles :'It is a psychological fact that we believe we have the ability to control and modify our actions by the exercise of "will", and in practical life all sane men will assume they have this ability'. Nevertheless free will has a very unpopular history in science because of the paradox of mind acting on a supposedly deterministic physical system. The controversiality of consciousness and free-will in science stems from their supposed inconsistency with causal scientific description - but is this so? Chaotic dynamical systems and quantum mechanics both share attributes which are more consistent with these two features than biochemical reductionism would suggest:
1.2.1 Chaotic dynamics possess both sensitivity to initial conditions and computational unpredictability, fig 1(a) (Schuster 1986, Stewart 1989) because exponential spreading of adjacent trajectories with increasing time occurs with a positive Liapunov exponent L > 1 fig 1(a). Similar considerations apply to complex systems in transition in and out of chaotic regimes, where L crosses 1 . Arbitrarily small fluctuations are thus inflated into global instabilities by the "butterfly effect", in which a perturbation such as a butterfly can become the source of a subsequent tropical cyclone. A chaotic system is arbitrarily sensitive to perturbation and hence responsive. Sensitivity also prevents a chaotic system from being precisely modelled. It is thus unpredictable, though it may be deterministic. Such systems behave in a way which makes it impossible for an observer outside the system to describe it precisely enough to determine its outcome and hence test its causal nature.

Fig 1: (a) Positive Liapunov exponent causing sensitive dependence
on initial conditions is illustrated by the Lorentz flow. Trajectories starting
e-close at a have exponentially separated by b and are in distinct parts
of the flow by c1,2. Computational unpredictability follows from the incapacity
of any approximation to model the flow over increasing time. (b) Non-integer
fractal dimension D of the Koch flake. Each side is repeatedly replaced
by 4 sides of length 1/3. The Lorentz flow has a strange attractor which
has a correlation dimension of 2.06 (Grassberger & Procaccia 1983).
1.2.2 Quanta possess a global internal model of their environment in the form of the wave function and violate causality through uncertainty during reduction of the wave packet. These attributes, although somewhat different in nature from sensitivity and unpredictability, cap off the causality of chaotic physical systems at the microscopic level and have parallels to consciousness and free-will. Although a quantum behaves like a particle in terms of its emission and absorption, its wave function extends throughout space-time, forming a global representation of its environment. Subsequent absorption of the particle appears to collapse the wave function into only one of its superposition of possible states. Thus although averaged ensembles of events conform to the probability distribution of the wave, the absorption foci of individual quanta cannot be predicted. The stochastic nature of the theory makes it possible in effect for each quantum to both be conscious of the universe through the extended wave function and to exercise free-will in its demise in wave packet reduction, while conforming to the principles of quantum mechanics.
1.2.3 Fractal linkage. These two levels of physics have a natural link of scale through the fractal nature of chaotic dynamics. Many chaotic systems display conserved sets such as attractors and basins which are fractal in the sense that they are replicatively self-similar on descending microscopic scales, resulting in a non-integer dimensionality (Peitgen & Richter 1986). In fig 1(b) the Koch flake illustrates a simple geometrical fractal in which each side is repeatedly replaced by 4 sides of 1/3 the length l. The number of units thus scales as l -log4/log3, giving a fractal dimension D = log4/log3. The correlation dimension is a similar measure (Grassberger & Procaccia 1983).
The term quantum chaos has been used to describe a variety of quantum systems which have analogous dynamics to classically chaotic systems. Electrons, atoms and small molecules traversing a molecular milieu, have dynamics well approximated by a chaotic system. Complementing this frontier is the link between quantum uncertainty and sensitive dependence. In a chaotic physical system, quantum fluctuations will become amplified into global instabilities by the butterfly effect - quantum inflation. The fractal nature of chaotic dynamics results in a disseminated set of increasingly unstable regimes which can serve as nuclei for global fluctuation (King 1991). Complex systems in the chaotic transition region may also enter a state of self-organized criticality in which a local perturbation leads to global bifurcation.
Although our sensory experiences, particularly visual representations of the physical world, have obvious stability properties, the necessity of sensitive dependence on the external world guarantees that the internal model must include unstable dynamics, as is confirmed by study of optical illusions, hallucinations and dreams. Recent studies of the chaotic dynamics of a variety of neurophysiological processes support such attractor-bifurcation models at both the cellular and neurosystems level.
1.2.4 The Mind of the Uncertain Brain Such a linkage could be used by the brain to allows mind states to critically perturb brain states through quantum non-locality. Such an interaction is possible only if the time evolution of the brain is formally unpredictable when observed externally as a physical system. This unpredictability must also be advantageous in evolutionary terms, and hence must assist the brain in the computationally intractable task of survival in the open environment. In this sense, mind may be an extra-physical dual aspect of reality, corresponding to unpredictabilities and uncertainties in a physical brain. Such an interpretation is a dual model of reality in which mental and physical are fundamental components of the universe. The physical world is manifest only indirectly as a set of stability properties of mind. From the physical perspective the same mental experiences correspond to stability structures in an internal model of reality.
2 Conceptual Problems in the Mind-Brain Relationship
2.1 Defining Mind and Consciousness
2.1.1 Mind and Consciousness remain so different from the usual objects of scientific inquiry that it is unclear they conform to the objective criteria usually applied to scientific description. The terms are the subject of such ambiguity that it is essential to review their semantic variations:
Mind is often referred to as the sum of our intellectual faculties, our capacity to think or reason, the rational mind, as distinct from sensation. Being out of one's mind in a complementary way suggests escape from rational controls into insanity. Mindless activity similarly implies unreflecting action. Minding carries different emphases of attending - "Mind that step!" or caring - "I don't mind if I do!". Being mindful again implies attentiveness. Having something on one's mind that of preoccupied thought. Making up one's mind that of deciding. Being of one mind having the same opinion or intent. Nevertheless, mind as the dual of body embraces the diversity of subjective experience. To avoid such semantic confusion, I will inclusively define mind as: the envelope of all subjective manifestations of brain-related activity, including sensation, thought, feeling and states such as dreaming. The essential feature of the mental is its fundamentality, the fact that all our access to the physical world occurs via the conduit of subjective experience, the one undeniable reality of existence.
Two scientifically controversial attributes of mind are likely to be pivotal to a successful description :
2.1.1.1 Consciousness : subjective manifestation of attentive brain function.
2.1.1.2 Free-will : mental action upon brain function, violating physical determinism.
At a primary level, our conscious experiences are our only access to the nature of physical reality. Although we become firmly convinced of the existence of the real world, our access to this world is only by inference from our subjective, conscious experience as observers. The physical universe is manifest entirely as structural stabilities of conscious experience, or as conceptual descriptions we develop from analysis of other forms of conscious observation, such as particle tracks, spectrographs etc. The physical world is thus accessed indirectly as an inferential structure linking our own conscious experiences and those of others into a single stable description. Thus despite believing in the primacy of the world we live in, all of us derive all our knowledge of the universe from our subjective representations of reality (Eccles 1966, Blakemore & Greenfield, 1987, Rose 1973, Margenau 1984). Without the direct avenue of subjective awareness, it is unclear that a physical universe would even exist.
This basic and undeniable manifestation of subjective consciousness has again a backdrop of other more restrictive concepts. One can refer to another person as conscious or unconscious depending on whether they appear to be responding to stimuli, or in a state of coma. One can also compare the conscious with the subconscious or the collective unconscious of Jung, allowing for some attributes of mind to pass subliminally into existence or represent archetypes lying below the level of attentive awareness. Cognitive scientists may dismiss many peripheral aspects of brain processing as being pre-conscious, leaving only the major orienting focus of attention as reaching the conscious level. Finally consciousness is viewed as a reflective type of attention process, exemplified by self-consciousness in which a person in addition to perceiving is aware of themselves as a subject. As described by Rosenthal (1986) "a conscious state is one which has a higher-order accompanying thought which is about the state in question". Ironically such self-consciousness has been claimed by Jaynes (1976) to be a very recent and culture-born imposition, however to the contrary, it may be an evolutionary feature of mammalian or even metazoan brains, because distinction of self from world is the fundamental distinction for survival. A central feature of consciousness research is how perception and attention combine to result in action in the brain (and mind).
I will thus inclusively define consciousness as: the envelope of capacities of the brain to form subjective representations of reality. In this sense, the brain is conscious of mind. This is consistent with one of the most popular modern conceptions of the mind as an internal model of reality constructed by the brain, a view consistent with a variety of lines of evidence spanning normal cognition, the nature of optical illusions, dreaming and many pathological states of the brain. Free-will on the other hand is the reciprocal action of mind on brain.
2.1.2 The Menagerie of the Mind The scope of subjective experience can be outlined through a combination of introspection and study of traditional concepts in language. The most undeniable mental experiences are our sensory experiences of the world around us, and physical sensations in our bodies from touch and pain through to the manifestations of emotions such as fear and anger. Along with these come less clearly defined inner images provided by memorization, imagination and thought. While these are less rich than direct sensory experience in usual waking life, probably as a result of competition with direct sensory stimuli accessing the same processing modules, in states of relaxation, sensory-deprival, dreaming, or meditation, internal imagery becomes clearer and less distinguishable in intensity from sensory experience, and may take on a life of its own. Study of the properties of such alternative states has immense potential for consciousness research, because the properties of consciousness are not sensorily tied to the physical world description, thus providing a unique opportunity to test its internal dynamical qualities.
Many mental features, being processes rather than states, cannot be subjected to stable scrutiny without changing their characteristics. While it is easy to understand the meaning of this sentence, it is vastly more difficult to simultaneously perceive the details of how you are able to understand its meaning, and more difficult still to decide to what extent the semantic background of the words is consciously perceived or merely subconsciously controlling the conceptual process. Nevertheless it is accepted English usage that we can choose to think and that thinking is a structured conscious process, in which a sequence of semantic concepts emerge as a product of directed voluntary attention, a sequential process with semantic direction, which may take the form of narrative, rehearsal or have sensory or abstract properties. To imagine is again a voluntary process but dealing with imagery generation. By contrast, to have an idea is a spontaneous and possibly novel product of the thought process. Such terminology is consistent with a dynamical model in which the attention process generates structural bifurcations, some of which spontaneously generate novel features or concepts. The difficulty of attending to two things at once and the singularity of the stream of consciousness suggests that it represents a global dynamic in the brain. It may be more accurately described as a bundle of related awarenesses, dynamically coupled, but capable of further bifurcation.
Central to our subjective consciousness is the notion of an observer, or self, witnessing a theatre of conscious experience through the attention process, having the capacity by an act of will, or intent, to alter the circumstances occurring in the perceived world. Dennet (1991), based on the dictates of a parallel processing model, and various perceptual illusions, asserts that such a "Cartesian Theatre" is non-existent. "What would make this sequence the stream of consciousness? There is no one inside , looking at the wide screen show displayed all over the cortex, even if such a show is discernible by outside observers". Nevertheless the construction of the physical world model is derived from our conscious experiences. While there may not be a localized and distinct representation of the stream of consciousness in the brain, and consciousness, including time-perception, may present as an internal model of reality, the self and its intentional relation to the stream of consciousness remains our fundamental arena of experiential reality.
2.1.3 Observer Problems in Consciousness Several features make consciousness uniquely difficult to observe:
2.1.3.1 Mental observation [introspection] involves a whole-systems observer problem in which the system being observed is also the system doing the observing. "Introspection is one of the many forms consciousness itself can assume, so that it represents a significant part of what we are trying to explain" (Miller 1992). The attempt to examine conscious states thus globally alters their phenomenology.
2.1.3.2 Mental constructs do not satisfy the criteria of objectivity possessed by physical objects and processes, so most aspects objective observation are rendered invalid. "Although consciousness exists by virtue of some physical property of the brain, just as bioluminescence exists by virtue of some chemical property of specialized cells, it is not as bioluminescence is an observable property of living matter. Nor is it an invisible property... It is detectible to anyone who has it. The difficulty is the method by which consciousness is detected is logically different from ... bioluminescence" (Miller 1992). Thus qualia such as redness remain ill-defined.
2.1.3.3 As William James originally pointed out "consciousness is not a thing, but a process". Conscious features may thus be rapidly transformed by the very act of observation, leaving only the sensoria as stable observables. Thought is an example of such dynamically unstable observation, because the act of thinking results in an evolution of the experience. An idea is an unstable bifurcation to a novel thought state.
2.1.3.4 The boundary between subconscious processing and conscious experience is ill-defined. Consciousness may thus be incomplete as a process. The hidden background of conscious experience, which Dreyfus and Searle call "intrinsic intentionality" may contain some of the highest computations.
2.1.3.5 Crick & Koch (1992) note that the problem of consciousness is ill posed, because it is a constructive process, requiring further constraints to provide a unique solution.
2.1.3.6 The subjective experience of free-will is inconsistent with causal description.
2.2 Concepts of the Brain-Mind Relationship
A diverse variety of views of the mind-brain relationship have been advanced, providing pointers to the basis of mind in brain function, while confirming the unresolved state of understanding in this area.
2.2.1 Complexity : Consciousness as a property of complex computational systems. One common idea about the emergence of mind is that it is somehow a product of the very complexity of the brain. However neither computer circuits nor artificial neural nets are ascribed to possess the attribute of mind in addition to their functioning at any given level of numerical complexity.
John Searle (1980) invented the Chinese room, in which [conscious] operators translate Chinese by symbolic constructs without the system being conscious of the meaning of the translation, to point out that strong AI cannot by itself explain consciousness. Dennett (1991) utters the 'systems reply' against this by saying that the brain is complex and is conscious so it is possible in principle for a parallel virtual machine to become conscious through it's many parallel interactions - "this little bit of brain activity doesn't understand Chinese, and neither does this bigger bit of which it is a part ... even the activity of the whole brain is insufficient to account for understanding Chinese"... of course the dualists would say "it takes an immaterial soul to pull off the miracle of understanding". Dennet's critical untested assumption is that the brain is simply a classical virtual machine.
Measures of complexity based only on neural net architecture neglect non-trivial properties of the individual neuron. The coelenterate Hydra, fig 2, displays complex behaviour involving a greater variety of locomotion types than the sea snail Aplysia and coordinated feeding, despite having an undifferentiated nerve net, which can withstand migratory reintegrations of the ectoderm and endoderm if the organism is turned inside out.
Certain non-linear systems near the chaotic boundary L ~ 1 or in chaotic transition do however display complex bifurcations and the development of novel structure within the system, and for this reason are termed complex systems by the Santa Fe school (Jen 1990). These form good models for neurophysiology, and have the potential when given an adequate quantum foundation also to explain consciousness.

Fig 2: Hydra poses a dilemma for theories of neural net organization.
It can reassemble ectoderm and endoderm if turned inside out and has a disseminated
neural net (a) with no global structure, nevertheless it can coordinate
eating (b) and possesses more diverse types of locomotion than animals such
as molluscs and arthropods which have structured ganglia. These include
snail-like sliding, tumbling, inch-worm motion and use of bubbles and surface
films.
2.2.2 Emergence: The idea that new properties emerge from the interaction of subsystems, not predicted by the subsystem architecture. In this sense emergence, which is a property of many digital and non-linear systems including molecular assemblies, applies to virtually every system with global feedback, from machines and organisms to the weather, all of which share system attributes which are disrupted by partitioning. Thus while the basis of mind is doubtless also emergent, this does not in itself explain how it 'emerges' from the brain. It is thus necessary to look in detail at the underlying mechanisms of mind for a substantive explanation.
2.2.3 An internal model of reality constructed by the brain. The idea of the mind as an internal model of reality has widespread support, both in the modular structures of sensory detectors which respond to features such as line orientation, thus synthesizing a model out of a combined set of features, and in various visual illusions in which these features lead to incomplete or contradictory results. The internal model is also consistent with a role for the mind as a monitoring system for attention. However the precise nature of the model and how the brain uses it to generate mind remain to be elucidated.
On a physiological level, research into modular sensory processing (see 3.3) and the distributed chaos models of Freeman's group (see 4.1.1) illustrate internal models of brain function which share distributed processing features. At an opposite extreme Blackmore's (1988) internal model unites a variety of modes of mental experience surrounding dreaming into a common description.
Somewhere in between stand the purely structural models of artificial intelligence (AI) and cognitive psychology, which draw from the studies of distributed processing in modern physiology (see 3.1) to present parallel cybernetic models which, although they have powerful features, explain consciousness away as an elaborate form of parallel computational control of the organism. Dennet's (1991) multiple drafts model is an example which denies our subjective view of a theatre of conscious experience with an attentive observer. "There is no single definitive "Stream of Consciousness" because there is no Central Headquarters, no Cartesian Theatre where "it all comes together", for the perusal of the Central Meaner. Instead of such a single stream (however wide), there are multiple channels in which specialist circuits try, in parallel pandemoniums, to do their various things, creating Multiple Drafts as they go. Most of these fragmentary drafts of "narrative" play short-lived roles in the modulation of current activity but some get promoted to further functional roles, in swift succession, by the activity of a virtual machine in the brain. The seriality of this machine (its 'von Neumanesque' character) is not a hard-wired design feature, but rather the upshot of a succession of coalitions of these specialists", (see also Marcel & Bisiach 1988, Baars 1988).
The multiple drafts model makes some physiological sense because an asynchronous parallel architecture allows the brain to make optimally rapid, but arbitrarily complex calculations. Such an architecture is consistent with cortical design in which the incoming afferents are associatively connected with the outgoing efferents with no intervening structures, leading to very short computational delays, while the timing delays across the cortex are non-zero. It is also consistent with the attractor dynamical model advanced in this paper in which multiple bifurcations can induce parallel structures in the same dynamic. The general nature of such a distributed internal model also has very interesting time properties (see 8.2), and allows for constructive filling in [blind spot and fig 5(c)], editing, and for gaps in the description, such as those caused by visual saccades to be smoothed out (Dennett 1991) . "Nothing can seem jerky except what is represented as jerky" - Minsky.
However Dennett's description clearly leaves mind a role equivalent only to an abstract computational process. "The neuroscientists are right to insist that you don't really have a good model of consciousness until you solve the problem of where it fits in the brain, but the cognitive scientists, (the AIers ad the cognitive psychologists, for instance) are right to insist that you don't really have a good model of consciousness until you solve the problem of what functions it performs and how it performs them - mechanically, without the benefit of Mind. ... Anyone or anything that has such a virtual machine as its control system is conscious in the fullest sense and conscious because it has such a virtual machine" (Dennett 1991). "Any scientific theory of mind has to treat it as an automaton" (Johnson-Laird 1983). These statements lose the subjective nature of mind completely.
This difficulty arises from dependence on abstract computational ideas as the basis for the models, reducing the functional implementation, and particularly its neurophysiological basis to subordinate roles as in Marr's (1982) three-level analysis of mental phenomena. If the brain uses unusual physical principles in its neurophysiology, such models will fail by the classical trap. Repeated objections have also been mounted on philosophical grounds. "No connectionist computer could exhibit real mentality" (Searle 1990a,b). "The subjective features of conscious mental processes - as opposed to their physical causes and effects - cannot be captured by the purified form of thought suitable for dealing with the physical world that underlies the appearances" (Nagel 1986). The current paper addresses this problem by presenting a model in which the biophysical principles are paramount and the computational features are a product of the biophysics.
2.2.4 Duality : Mind as a dual phenomenon to physical reality. The very great difference between mind and the diversity of physical phenomena suggests that although in a sense paralleling the physical brain, mind may not be a physical attribute as such but a complementary principle, making mind and universe dual aspects of totality. Descartes' homunculus in the pineal and his "cogito ergo sum" illustrate two aspects of his dualism, expressed very eloquently in the following passage: "While I could pretend that I had no body, that there was no world ... I could not pretend that I was not ... from the fact that I thought of doubting the truth of other things ... it followed I existed ... from this I recognised that I was a substance whose whole essence or nature is to think and whose being requires no place and depends on no material things" - Descartes Discourse on Method 1637.
Dualism has been the subject of attack and counter-attack during the 20th century. Gilbert Ryle (1949) made a classic critique of dualism, describing it as the "ghost in the machine", and branding many aspects of mental description as category mistakes, promoting Koestler (1967) to mount a rebuttal by the same title. An outstanding feature of many of Ryle's descriptions, such as the lemon on the desk, is that they derive from everyday experiences in which the stable physical world model dominates other aspects of conscious perception.
Several researchers have proposed dualistic philosophies or even three-aspect brain-mind-knowledge models. Sir John Eccles wrote "we are a combination of two things or entities : our brains on the one hand; and our conscious selves on the other." The brain is a precious "instrument" a "lifelong servant and companion" providing "lines of communication from and to the material world," but we are not it. An act of will, as Eccles sees it is an everyday case of psychokinesis of mind moving bits of matter (Hooper & Teresi 1986). He has even gone as far as to propose structured entities psychons corresponding to ideas which interact with dendrons possibly in the pyramidal tracts. Vendler (1972, 1984) has also made a defence of dualism. Epiphenomenalism constitutes a weak one-sided form of duality in which world effects mind but not vice versa.
Critics of the artificial intelligence and cognitive psychology schools such as Dennet (1991) describe dualism as forlorn because it undermines the agenda of materialist or operationalist description of both the brain and mind in abstract computational terms ... "If mind and body are distinct things or substances, they nevertheless must interact; the bodily sense organs via the brain must inform the mind, ... and then the mind, having thought things over must direct the body in appropriate action. ... Cartesian interactionism". "How can mind stuff both elude all physical measurement and control the body?". Such criticisms depend on outmoded classical reasoning ... "It is [the] principle of conservation of energy that accounts for the physical impossibility of perpetual motion machines, and the same principle is apparently violated by dualism. This confrontation between quite standard physics and dualism has been endlessly discussed since Descartes's own day, and is widely regarded as the inescapable and fatal flaw of dualism". This statement displays a startling lack of understanding of a variety of physical processes from the weather to the behaviour of quanta, in which instability and uncertainty fragment classical causality and provide room for a logical loop connecting mind and brain - a central theme of the current paper.

Fig 3: (a) Nor-epinephrine and serotonin pathways originating
from mid-brain centres modulating light and dreaming sleep are distributed
across wide areas of the cortex (Bloom et. al. 1985). (b) Inputs from different
areas impinge on a singe hippocampal CA1 neuron via distinct neurotransmitters,
thus mapping neurosystems architecture on to single cells (Alkon 1989).
(c) Looping circuits of the limbic system are involved in both affect and
memory. (d) Limbic structures and the thalamus in brain section (Bloom et.
al. 1985).
2.2.5 Sensitive instability. The internal model is non-linear, dynamically unstable and undergoes transitions to and from chaos. While some aspects of mind, such as visual processing, have structural stability in their representation of most objects, optical illusions and a variety of mental phenomena from hallucinations to the bizarre subjective realities of dreaming sleep demonstrate that the mind is capable of unstable self-generative behavior. Dynamical arguments dictate that a nervous system which is optimally responsive to sensory conditions must undergo transitions into chaos to provide arbitrary sensitivity and ensure it does not become locked in any stability state or periodic oscillation. Thus while philosophers such as Gilbert Ryle, who rely on stable mental representations of objects, may conclude that mind is not even a well formulated concept, the unstable aspects may be essential to the internal model of reality.
Sensitive dependence is consistent both with representation of the mind as an internal model and with a dualistic interpretation of reality. The form of stability sets such as attractors and their evolution through bifurcation provides the basis for an internal model. The unpredictability of the chaotic regime and its ramifications in terms of quantum uncertainty provide a causal loophole within which to discuss principles complementary to physical processes occurring above the quantum level, which would support the dualistic perspective without contradiction.
3 The Neurophysiological Basis of Mind
It is obvious that the manifestation of consciousness in the forms we are used to experiencing requires interactive dynamics of the sensitively dependent active brain. What remains more elusive, like the engram of memory, is the neurophysiological basis for conscious states.
3.1 Local versus distributed generation of mind.
The hypothesis that mind is a consequence of the nature of distributed brain processing has a variety of support, based on the modular architecture of the cortex, modal arousal of the ascending distributed pathways and the impact of limbic circuits which we will investigate below. As all these systems are required for conscious attentiveness, the conclusion that consciousness is a global manifestation of attentive brain function is strongly supported.
An alternative strategy for the brain to generate the mind would be to have specialized areas dedicated to the generation of the internal model of reality. Sir John Eccles suggested that the mind is generated by the supplementary motor cortex, which is almost universally active (Popper & Eccles 1977). However damage to a variety of cortical areas reveals only "graceful degradation" of both function and subjective awareness, rather than 'mind loss' (Blakemore 1991).
Disagreement over the source of major oscillations in the brain such as the alpha and theta rhythms has led to ideas that specific sub-cortical structures such as the thalamus may play a principal role in generating mind, acting as a gating mechanism between the cortex and subcortical sensory centres (Crick 1984, Taylor 1992). It has topographical representations of many cortical modalities and forms an intermediate position between the cortex and the sensoria, thus having the potential to act as a filter releasing only the most relevant stimuli into conscious attention, coinciding with Huxley's (1954) filter theory of consciousness. These roles remain to be proven because the thalamus is difficult to access and the evidence from accidental lesions in humans is not clear. Similarly the ascending pathways of the basal brain are essential to maintain active consciousness, but this appears to be by diffusely activating the entire cortex.
Consciousness could nevertheless involve only a subset of cortical neurons. "Consciousness corresponds to coalitions of cortical neurons dynamically modulated by attention and higher-level expectations ... We would expect that only some of the cells in the cortex would be expressing what we see, while others are carrying out unconscious computations, but which cells correspond to consciousness?" (Crick & Koch 1992). At an extreme, identifying mind with distributed processing becomes mind-brain identity in which all mental states are considered the subjective equivalent of particular brain states. Mind-brain identity has two difficulties. Firstly the structural basis of mind is eliminated rather than explained by identity. Any responsive physical system, such as a computer could equally possess mind by the identity principle. Secondly identity prevents any causal link between the mind and brain, reducing consciousness and free-will to the status of delusions of the internal model, eliminating any role for the subjective aspect of reality in evolution and hence the need for an 'identity' mind.
3.2 Ascending distributed pathways. A very important contribution to the modal nature of conscious activity comes from the distributed pathways ascending from the midbrain centres, fig 3(a) responsible for general arousal in the reticular activating system [RES], and in regulating the major modality shifts of consciousness, light and dreaming sleep. Two pathways lead from the Raphe Nuclei and the Locus Coeruleus to diverse cortical areas and involve the modulating neurotransmitters, serotonin and nor-epinephrine, fig 11. The onset of dreaming sleep is heralded by activity of cells in the Pons and silencing of cells in the Raphe Nuclei and Locus Coeruleus (Bloom et. al. 1986). The latter also show bursts during orientation to novel stimuli. Similar dopamine paths spread out from the Substantia Nigra selectively into the frontal lobes and motor centres. The ascending pathways have been implicated in mental illness, addiction and motor syndromes such as Parkinson's disease (Gilling & Brightwell 1982). Dopamine is sometimes associated with pleasure and nor-adrenaline with anxiety.
Dreaming or REM sleep is both one of the most singular phases of conscious activity in which feedback appears to be accentuated at the expense of external input, generating complete subjective realities or 'worlds within'. The nature and function of dreaming consciousness and its wealth of detail remains obscure. The principal hallucinogens are also serotonin and catecholamine analogues, fig 11. In addition to cerebral arousal and sleep loss, their action probably triggers fractal waves of cortical excitation through the diffuse connections into interleaved cortical layers shown in fig 3(a) inset. Such connectivity gives the hallucinogens the capacity to modulate reflex modular excitability across the entire cortex, giving rise to patterned visions and cross-sensory synesthesias. They have earned a controversial reputation by provoking, alongside dreaming, some of the most remarkable changes in the nature of consciousness discovered by man. For the same reason they represent a key tool for consciousness research.
3.3 Cortical modularity. A variety of experiments support the modular (Fodor 1983, Mountcastle 1978) involvement of cortical centres, both in various sensory-cognitive tasks and their correlates in mental activity. These illustrate how modular cortical processing can be associated with different aspects of conscious experience.
Visual processing is clearly modular, with parallel processing of colour and movement in distinct areas (Zeki 1992). The primary visual area V1 has 'discrete and segregated' connections through more structured sensors in V2 to V3, V4 and V5 responsible for dynamic form, colour form and motion. These areas are each associated with their own lesion syndromes such as achromatopsia [no colour], akinetopsia [no movement]. Zeki's studies raise fascinating questions concerning how subjective visual awareness is generated from these modular aspects. Although they have direct reciprocal connections and diffuse connections back to V2 and V1, there is no central arena where their output is sent, suggesting visual awareness is generated either by their modulation back on V1, or somehow by the entire distributed dynamic. Face and mood recognition, written and spoken speech, and music, are all modular features.
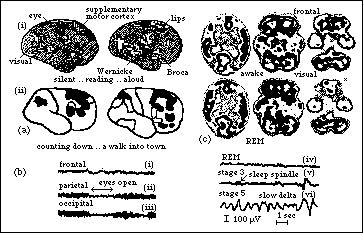
Fig 4: (a) Cortical modularity in PET scans. (i) Silent reading
is contrasted with reading aloud, in which there is wider cortical involvement,
including motor and somatosensory lips areas. (ii) Counting down is contrasted
with visualizing a walk into town, stimulating a variety of frontal areas
as well as the parietal cortex (P. Roland ex Blakemore 1991). The supplementary
motor cortex is activated in all four views. (b) Similarity of the REM sleep
EEG (iv) to the alert state [(i)-(iii) eyes open]. (c) REM sleep PET scan
parallels the awake brain in activation of visual cortex and frontal lobes
[dark], except for reduced REM inferior frontal activity (*), consistent
with the lack of control in dream sequences.
The PET scans in fig 4(a) illustrate the correspondence between different conscious activities and the activation of specific modular regions of the cortex, including well-known areas such as Wernicke's and Broca's areas of linguistic interpretation and articulation and the supplementary motor cortex, as well as highlighting less well understood areas in the frontal lobes. Listening to ambiguous structured signals [time reversed recordings] can activate just about the entire cortex (Friberg 1992) suggesting that activation is at least partially a product of chaotic instabilities in the process of forming a stable representation. Dreaming states provide interesting PET scans in which the cortex is perceptually activated in a manner consistent with the similar EEG to the waking state, fig 4(b,c), and the intensity of dream experiences. One feature different in dreams is the lower activity of the inferior frontal cortex, which may reflect the uncontrolled nature of dreams (Madsen et.al. 1991).
Such studies support a modular distributed internal model, which may also be holographic in its use of distributed phase fronts and wave transforms. The holographic aspect, popularized by Pribram, arises naturally from the many to many nature of synaptic connections, the simultaneity of Hebbian synaptic response and from the decomposition of sensoria into features such as line orientation and ocular dominance, forming a parallel representation in which each memory or experience is registered across the cortex as in terms of its attributes. What is less clear is how attributes such as colour motion, edges and filled regions coordinate with the primary sensory areas to resynthesize the complete conscious view.
3.4 The Split-brain and sex. Study of split-brain patients reveals how difficult it is to pinpoint the structures supporting sentient consciousness. When the left and right cerebral hemispheres are severed by cutting the corpus callosum, replies to experimental questions centre around the content of the dominant hemisphere, because it is uniquely able to articulate verbal responses. If a composite image is shown separately to the left and right half eye fields of the split-brain subject, they will describe the image their dominant hemisphere sees, and will be unable to refer descriptively to stimuli presented exclusively to their recessive hemisphere, unless it also retains linguistic ability. For example a nude in the left field will elicit a laugh and the indirect comment "That funny machine". The person thus behaves verbally, but not emotionally, as if the experiences of the non-dominant hemisphere are unavailable. However if asked merely to point to the picture seen, the recessive hemisphere, which is specialized at non-verbal pattern recognition, can assume control. The subject appears to retain an integrated personality and displays only minor conflicts of behavior, because the midbrain structures involving emotion and arousal are still intact, although the hemispheres appear a little like Siamese twin minds which can express distinct desires and aims. One patient's hemispheres for instance had the distinct aims of being draftsman and racing driver (Gazzaniga in Bloom et. al. 1985). Consciousness, even in the intact brain, may be a bundle of coupled awarenesses (Gazzaniga 1985, Parfitt 1987, Dennett 1991) rather than a single dynamical entity.
Sexual differences in the brain provide a further intriguing dimension of variation in human consciousness (Kimura 1992). At least three types of difference have been noted, enlargement of a specific hypothalamic nucleus in men, and differences both in lateralization and relative function of frontal and parietal areas. Women appear to have less linguistic lateralization, despite displaying superior verbal abilities (McGlone in Bloom et. al. 1985), and are several times more likely to have aphasia or apraxia from frontal than parietal strokes. Men have corresponding parietal and lateral dominance, leading to the possibility that male and female brains have global differences of organization similar to the plasticity seen in sensory processing.
3.5 Attention and Memory : The Limbic System and Frontal Areas.
The limbic system fig 3(e), (Mishkin et.al.1988, Alkon 1989) also forms a particularly interesting set of looping pathways combining sensoria, emotional states and episodic long-term memory, (c,d). The hippocampus, which has an older three-layered structure than the six-layer neocortex has a pivotal role in establishing long-term memory, possibly by transferring experiences into associative memory over a matter of weeks. It has projections from diverse sensory areas via the entorhinal cortex and feeds back into the thalamus and subsequently to cortical areas including prefrontal, cingulate gyrus, and basal forebrain. The amygdala has similar looping circuits linking diverse sense modes and connecting to the thalamus and deeper emotional centres in the hypothalamus. Phase decoherence occurs in the hippocampus during orientation to unfamiliar stimuli. The limbic system thus forms a bridge between emotion, memory and a distributed representation of time action and self in the frontal lobes. This is consistent with the emotional and motivational side-effects of frontal lobotomy and with studies on time-delayed learning in damaged hippocampi. It also gives a graphic portrayal of the link between extremes of emotional experience from fear to exaltation, the central themes of survival, attention, and the establishment of long-term memory.
The limbic system may also be linked to both the ascending pathways and the cortex in processing waking memories in the REM phase of sleep (Winsen 1990). In several mammalian species, theta rhythms, which are generated in the brain stem and pass both to the Septum in the forebrain and to synchronous generators in the hippocampus, can act as phase locking signals for long-term potentiation, in association with orientation to key survival activities. Spatially selective CA1 cells, which signal an animal's location in the waking state, are selectively active in subsequent REM phases, suggesting REM functions in memory processing in the sleep cycle. The role of emotion in the limbic system as a monitor of and driving force for survival, may thus underlie the archetypal intensity of dream content. However, because we dream about past experiences, it is also likely that hippocampal cells will revisit locationary states. Notably, we can also remember our dreams. It is difficult to see how this is possible at the same time as memory reconsolidation.

Fig 5: Optical illusions illustrate the close linkage between
mind state and brain state, and support a dynamical bifurcation model of
conscious imaging. (a) The lower face, which is easy to recognise in either
orientation gives similar evoked potentials, while the upper one which is
difficult to recognise inverted does not (Jeffreys ex Blakemore 1991). Optical
illusions illustrate two features of visual processing consistent with attractor
dynamics: (b) alternating percepts of a diagram which is ambiguous in 3-D,
(c) construction of a geometrical figure from incomplete information.
A second active type of short-term memory often called working memory (Goldman-Rakic 1992) complements [long-term] associative memory by providing short-term storage of symbolic information as well as permitting the manipulation of that information. It appears to be centred on the prefrontal cortex with reciprocal connections with both the parietal cortex and the limbic system. Prefrontal damage effects use of knowledge to guide behaviour in everyday situations, including predictive tracking e.g. of projectile movements. When a monkey is trained to look at where a target has disappeared after a delay selected cells in the prefrontal cortex fire, on target disappearing, others fire during the delay and others on the motor act. Such eye movement also involves feedback loops to the basal brain [from pyramidal cells to the striatum, the substantia nigra, mediodorsal thalamus and back to pyramidal cells]. Prefrontal action is modular with spatial and compound attributes active in different loci. A model linking the frontal and limbic structures in a representation of space-time is developed in 8.2.
3.6 Global Dynamics : Mind States and Brain states.
Examination of the contributions to consciousness by the major dynamical structures of the brain confirms that consciousness is a collective product of the ascending pathways, the cortex and limbic system. The dynamical activation of the cortex by ascending distributed pathways appears essential for consciousness, while all sensory areas and most of the associative areas of the cortex can be seen to contribute in a modular manner to the envelope of conscious experience, dynamically modified by the looping circuits of the limbic system.
Conscious experience is thus subject to much of the global dynamical variation for which the brain is capable, from anaesthesia, through waking, resting, dreaming, and hallucination. Only in deep sleep, seizure and coma when the global dynamics are profoundly altered do we see a loss of conscious function. In fig 5(a) the close correspondence between conscious recognition and brain states is illustrated. In (b) two optical illusions illustrate the way in which perception displays both oscillatory compensations in the presence of ambiguous 3-D information, and attractor-like constructs which complete partial visual information with perceived lines and regions. This can be seen as a possible example of linkages between edge detectors in V2 and higher areas and is also consistent with an attractor-bifurcation basis for the internal model of reality.
Crick and Koch (1990,1992) have explored a variety of hypotheses as to where and how consciousness might be generated in the cortex. One structured hypothesis is that it is associated with a specific frequency mode. "We have suggested that one of the functions of consciousness is to present the result of various underlying computations and that this involves an attentional mechanism that temporarily binds the relevant neurons together by synchronising their spikes in 40Hz [35-70Hz] oscillations". They point out that distinct images could excite the same modules without confusion if they had synchronous oscillations. This would explain, for example, how you can briefly imagine something while looking at the world around you. They also speculate that the top layers of the cortex could be unconscious and layers 5 & 6 which output to other areas might be conscious. A minimum processing time of 60-70ms for a conscious percept can be deduced from the display of rapid images [20ms red + 20ms green = yellow] or tones.
4 Non-linear Dynamics in the Central Nervous System
Physical evidence for attractor dynamics in a variety of aspects of central nervous system function and development has become abundant in recent years. Chaotic regimes have been demonstrated, both at the neurosystems level and in the dynamics of single excitable cells, and fractal dynamics has been proposed for ion channels (King 1991).

Fig 6: Chaotic neurosystem dynamics are illustrated (a) By
the low correlation dimensions of a variety of brain states, (b) phase portrait
of an EEG recording (Babloyantz & Salazar 1985), (d) time evolving Fourier
spectrum and associated correlation dimensions. (c) Phase decoherence in
novel or unexpected stimuli is consistent with a distributed model of processing
based on oscillatory phase, similar to a hologram. (e,f) Chaotic and distributed
processing is combined in Walter Freeman's model of olfactory bulb processing.
Recognition of a given odour by the bulb arises from the distributed pattern
of activity, occurring in bursts phased with inhalation. The time-dependent
dynamics supporting this consists of bifurcation from low-level chaos to
higher level activity which settles into an existing attractor [recognition]
or develops a new one [learning] after exploring phase space in higher level
chaos. Chaotic neural nets based on neuroanatomy can perform pattern discrimination
tasks competitively with other neural net designs (Skarda & Freeman
1987).
4.1.1 Neurosystems Dynamics: At the neurosystems level, experimental evidence has accumulated for chaos in the EEG's of at least some phases of cortical activity, including sleep, resting wakefulness and pathological states such as epilepsy (Babloyantz 1985, 1989, Basar 1990). The low correlation dimensions of several of these states is consistent with collective chaotic dynamics in neuron populations, rather than stochastic or independently programmed behaviour. Evoked potentials also show desynchronizations during orientation, consistent with phase relations being pivotal in recognition and orientation (Basar et.al. 1989, Hoke 1989), supporting a holographic model. The model of burst dynamics in the olfactory bulb advanced by Walter Freeman fig 6(e,f) combines these two aspects into a model with bifurcations of spatially distributed waves into chaotic temporal temporal dynamics which both follow the neurophysiology and also permit real tests of pattern discrimination of neural nets displaying comparable dynamics (Freeman & Baird 1987, Skarda & Freeman 1987, Freeman 1991, Yao et.al. 1991).
In this model, low level chaos is lifted into a higher energy state by olfactory input. Chaos in this state enables the system to explore its phase space, falling into an existing attractor in the case of a recognised odour, but bifurcating to form a new attractor in the case of a newly learned stimulus. The transition into chaos thus provides sensitive dependence on input, ergodic 'randomizing' phase space exploration, parametric bifurcation to form new symbols, and possible quantum amplification. Return from chaos in turn fixes stability structures from the fractal dynamics.

Fig 7: (a) Period 3 oscillations in Nitella indicate dynamics
within the chaotic regime. Chay-Rinzel model (1985), (b) output is comparable
with experimental recordings, (c) period doubling bifurcation in model.
(d) Experimental behaviour of voltage gated K+-channel is consistent with
a fractal model (3) rather than a one, or two step Markov chain (1,2) (Liebovitch
et. al. 1987a,b). (e) Fractal dynamics in relation of excited myoglobin
is illustrated in terms of functionally important movements and equilibrium
fluctuations involving a variety of quantum transitions on differing scales.
4.1.2 Single Cell Dynamics: Similar experimental evidence has accumulated for chaos in a variety of excitable cell types, supported by the chaotic models of Chay and Rinzel (1985), fig 7(a,b,c). These extend the Hodgkin-Huxley equations (1952) to take account of calcium ion pumping, thus displaying a variety of dynamical features, including period doubling bifurcations, chaotic regimes and period three oscillations characteristic of the chaotic regime. These dynamics model well the chaotic excitations of Nitella pancreas cells, and similar models have been advanced for excitations in neurons and heart pacemaker cells. The irregular behaviour of controlling cells in small ganglia such as in Aplysia studies is also consistent with sensitive dependence and chaos, by contrast with the more regular beating of subordinate neurons.
4.1.3 Fractal Ion-Channel Kinetics : A Markov model is commonly used for ion channel kinetics, in which an open state is linked to one or more closed states. However the likelihood that a closed channel will open behaves fractally with increasing time scales according to fig 7(d) (Liebovitch et. al. 1987a,b, 1991). This is consistent with bio-molecular structures behaving fractally, not only in a geometrical sense, but also dynamically as shown for myoglobin in (e) (Ansari et. al. 1985). The dynamics of many important biological molecules may be fractal in this way, which involves the linkage between a variety of quantum excitations of differing energies, and feedback between tertiary structures and active sites.
4.2 Long-term adaption. Many aspects of the long-term structure of sensory areas of the cortex are consistent with their inheriting an attractor structure derived from particular sensory innervation and stimulation patterns. On a short time scale are the reported spatial distributions corresponding to olfactory stimuli, fig 6(e) which on a longer time scale adopt new forms on relearning the same stimulus a second time, and phase desynchronization of cortical evoked potentials. On a longer time scale plasticity of sensory structure fig 7 from the somatosensory barrels of the rodent (b), through to visual ocular dominance and orientation patterns induced by training regimes (a) indicate major sensory structures arise as a dynamical system induced through bifurcation.
4.3 Neurogenesis. The raw numbers game of neurogenesis suggests attractor dynamics may form an essential bridge between central nervous genotype and phenotype. The 5 x 10^4 genes governing central nervous system development [around 60% of human genes] cannot informationally specify the connections for 10^11 neurons and 10^15 synapses. Generalizing principles are thus required to complement the informational specificity of developmental genes such as the homeotic genes, thus providing both genetic efficiency and plasticity. The form of early embryogenesis is a series of bifurcations induced by morphogen gradients leading to the differentiation of major brain structures, fig 9(a). Further general principles playing a dynamical role in central nervous organization include cell surface interactions enabling migration up the glial scaffold (b), selection of tissue layer, or target cell synapse type, and parallel growth (Blakemore 1991). The neurogenic picture is thus consistent with genetic selection up to target cell type but the development of specific synaptic connections and the organization of thalamic and cortical layers, ocular dominance columns, barrels etc. through dynamical excitation driven by the afferent sense organs, culminating in the sensory experiences of the newborn (Kalil 1989). This is consistent with the plasticity of sensory structures already described, with tissue culture experiments (c), and with the waves of excitation seen on the prenatal retina [1/10-1/100 action potential speed], which stop just before the onset of visual activity. These waves appear to result in both geniculate layering and subsequent organization in the visual cortex (Shats 1992), by competitive input, in which synchronous inputs cause Hebbian enhancement, while asynchronous inputs from distinct eyes cause synaptic separation, consistent with the sequential organization of (1) the retina via early firing in ganglion cells, (2) subsequent layering of the geniculate after retinal wave input, and finally (3) organization of ocular dominance and other features in the visual cortex.

Fig 8: Plasticity consistent with attractor dynamics is illustrated
(Blakemore 1991) (a) by field patterns of reversal of imbalanced ocular
dominance caused by covering the dominant eye in a cat, and (b) somatosensory
barrels in the mouse showing invasion by neighbouring nuclei on removal
of a whisker. (c) The visual field on the cortex is distorted by a non area-conserving
map similar to a complex logarithm, which gives greater sensitivity to the
centre but does not match the geometry of the perceived world (Cowan &
Sharp 1988). (d) Ocular dominance displays a bifurcation of the 2D field
in terms of the 1D parameter of dominance (Hubel & Wiesel 1989).